
Lec 1. 神经解剖学 Neuroanatomy
脑的外侧是颅骨、脑膜和幕骨,脑膜又分为三层。
外界传入大脑皮层经过丘脑,其中丘脑会将大部分信息阻止,只有少部分的信息达到大脑。
小脑是一个高度特化、反映迅速的器官,主导运动。在进化过程中倾向于主导程序化的、机械性的行为。这些行为是后天习得的;在早期的学习中,这些行为仍然由大脑皮层处理,并逐渐固化到小脑中,形成不经过思考的行为。
中枢神经系统:CNS,Central Nervous System
外周神经系统:PNS,Peripheral Nervous System
脑膜(Meninges)又分为硬脑膜、蛛网膜和软脑膜。硬脑膜提供机械性的支持和保护。
脑脊液有新陈代谢和缓冲的作用。脑积水是由脑脊液流动不畅引起的。
大脑皮层被沟回划分为四个主要的脑叶,外加上岛叶和边缘系统,一共分为:
-
额叶(frontal),主管认知。运动皮层(包括初级运动皮层和前运动皮层)位于额叶中,在中央沟(central sulcus)的前方,即中央前回(precentral gyrus)中。
-
颞叶(temporal),主管记忆。初级和次级听觉皮层位于颞叶,海马体位于颞叶的内侧。
-
顶叶(parietal),主管空间认知。顶叶涉及多种感知模态的整合,包括空间感觉、触觉和视觉的背侧流等。体感皮层(somatosensory cortex)位于顶叶中,中央沟的后方,即中央后回中。身体不同部分的感觉最终都会到达体感皮层中的相应位置,并形成侏儒人(cortical homunculus)的映射结构,见下图(为大脑自左向右的纵向切开面)。
-
枕叶(occipital),主管视觉。视觉皮层的大部分区域都位于枕叶,初级视觉皮层(primary visual cortex,V1)位于此处,腹侧流(ventral stream,空间通路,参与空间位置讯息和运动控制)和背侧流(dorsal stream,内容通路,参与物体识别)均从此处出发。
-
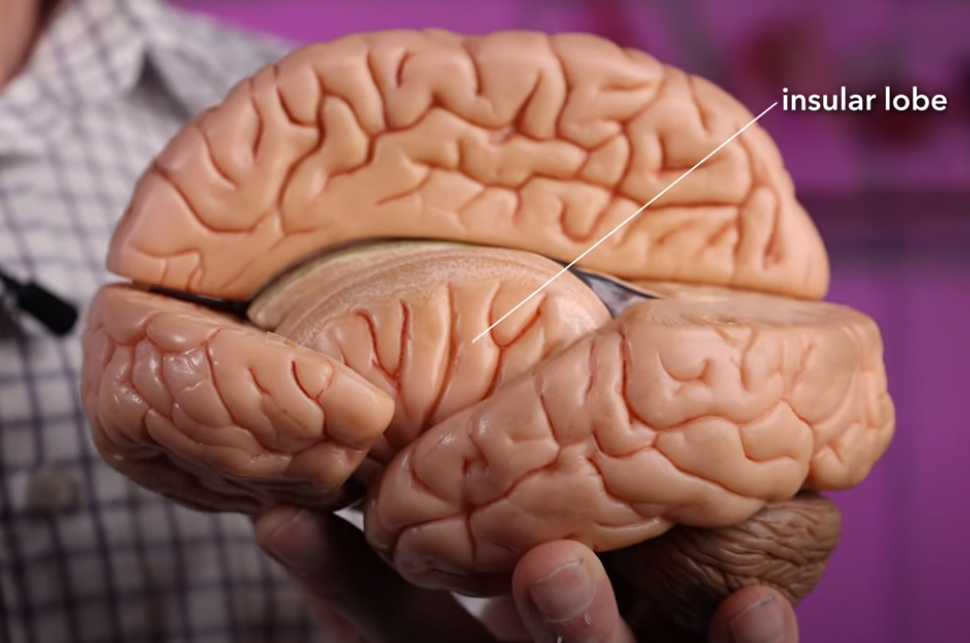
岛叶(insular),与身体感知和成瘾相关。岛叶无法在大脑的外部观察到,位于颞叶以内。
-
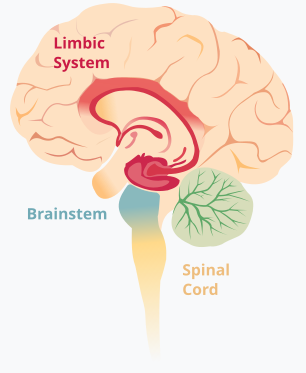
边缘系统(limbic),主管情绪。包含皮层以下的大量结构,海马体、杏仁体、丘脑等都被考虑为边缘系统的部分。
下图为侏儒人图像。侏儒人图像在运动皮层与体感皮层均存在,前者代表运动控制的映射,后者代表身体感知的映射。下图中,左侧为中央后回处的半脑截面,右侧为中央前回处的半脑截面。半脑截面代表对身体一侧的对应关系,另外半脑具有对称的图像,按照左脑控制右侧身体、右脑控制左侧身体的原则进行对称。
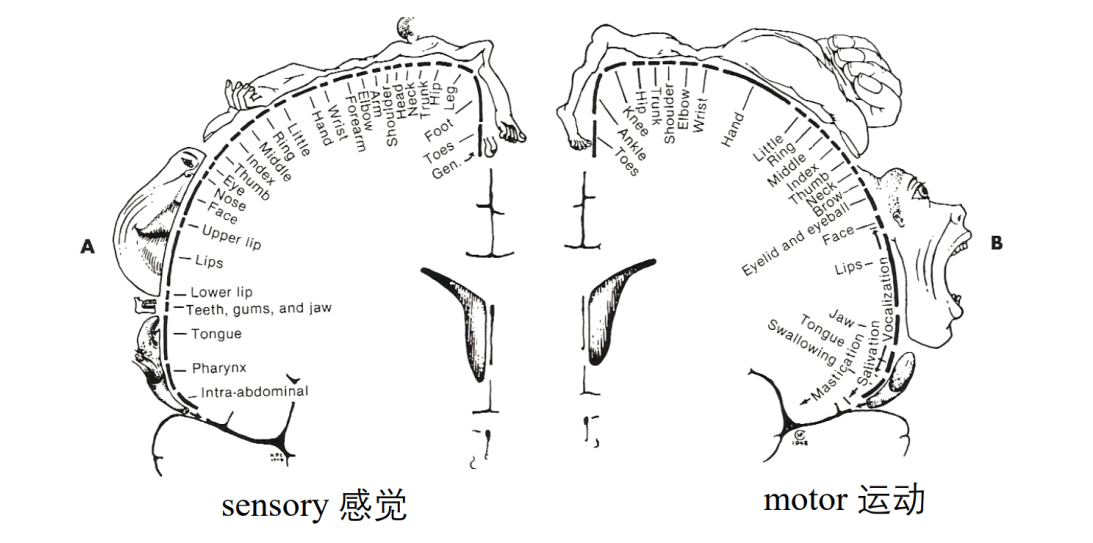
中央沟将运动区和基本感觉区分开。顶枕沟分隔顶叶和枕叶。
模块化在大脑中体现为功能柱。
Lec 2. 神经生理学 Neurophysiology
被动电学特性
简单的神经元特性,你知道的.jpg
膜电位外正内负,钾离子内高外低(-75mV),钠离子外高内低(+55mV)。平衡电势处于浓度作用和电势作用的平衡位置(-70mV)。Na+/K+ 浓度差由 Na+/K+ 泵维持,消耗一个 ATP 时,将 3 个 Na+ 转运到细胞外,同时将 2 个 K+ 转运到细胞内。
神经系统在轴突上通过髓鞘(myelin)包裹增加绝缘性,从而提高传导效率。髓鞘分段包裹,中继处由兰氏神经结(Node of Ranvier)进行电信号中继,即在相邻的兰氏结之间不断地去极化,形成新的动作电位,通过分段传播的方式提高电信号传导的速度,称为跳跃传播(saltatory conduction)。
动作电位
动作电位相对于分级电位/电紧张电位(graded potential),即阈值下动力特性,表现出不可叠加的特性。动作电位具有以下的性质:
-
或有或无。
-
阈值发放(-55mV)。
-
传导时不衰减,由兰氏结的传播方式决定。
-
不应期。
动作电位的发放由离子通道形成。当膜电位达到阈值时,Na+ 离子通道打开,Na+ 离子大量内流,形成脉冲。电位达到顶峰之后,Na+ 离子通道关闭,K+ 离子通道打开,K+ 离子大量外流,造成电位的下降和超极化。
突触与突触传递
电突触用于快速的信号传递和同步活动,可双向连接,通过神经元之间的缝隙连接(gap junction)形成。
化学突触略。
突触具有兴奋性或者抑制性。在兴奋性突触中,突触后递质受体在结合递质之后,打开通道,使得 Na+ 内流,从而引起电位升高。典型的递质包括乙酰胆碱和谷氨酸。在抑制性突触中,突触后打开通道时将引起 Cl- 内流,从而引起超极化。典型的递质包括-氨基丁酸(GABA)。
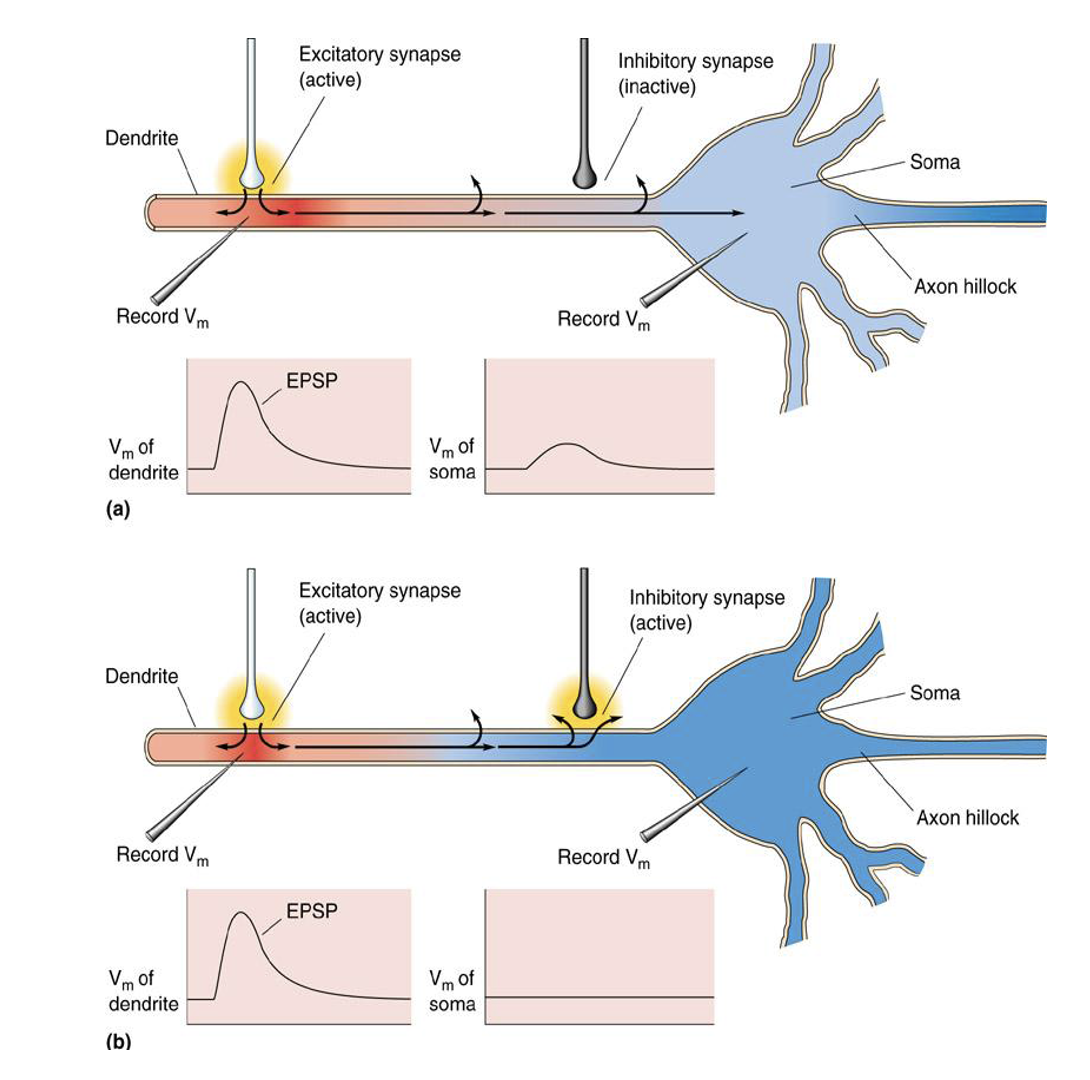
抑制性的神经信号整合还能够通过 shunting inhibition 完成。shunting inhibition 表现为阻止电信号在轴突上向胞体的传递,如下。
突触传递还会受到第二信使的作用。第二信使(second messenger)是一类在细胞内进行作用的物质,相对于第一信使。Ca2+ 离子是一种第二信使。第一信使指的是在细胞外起作用的物质,比如神经递质。神经递质可以通过与突触后膜上的受体结合,然后直接打开离子通道来影响信号传递,也可以通过第二信使产生间接的影响。一个典型的例子是 G 蛋白偶联受体参与的第二信使调控过程,如下:
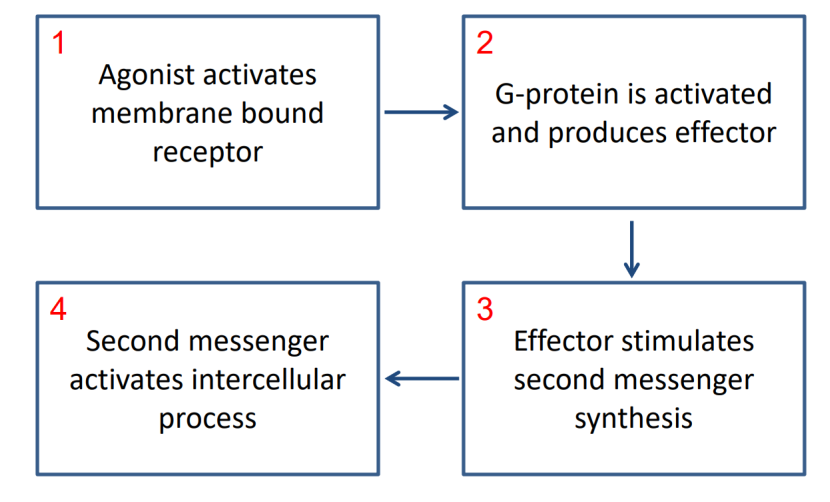
以肾上腺素激活肝细胞糖原分解为例,作用的过程是:
-
肾上腺素(epinephrine)作为激动剂,结合到肝细胞表面的-肾上腺素能受体(一种GPCR)
-
G蛋白的亚基交换 GDP 为 GTP,导致亚基与分离,激活的 G 蛋白亚基(Gs)激活腺苷酸环化酶(AC)。
-
腺苷酸环化酶催化 ATP 生成 cAMP(环磷酸腺苷,一种第二信使)。
-
第二信使在细胞内引起酶活化、基因表达调控、离子通道开放、细胞代谢变化等,从而引起细胞生化状态的变化。
第二信使系统具有信号放大作用。一个第一信使可触发成千上万个第二信使分子,进而激活大量下游蛋白,即级联放大作用(cascade)。这使得第二信使能够引起持续长时间的生化状态变化。
当物质满足以下条件时,它被考虑为一种神经递质:
-
在细胞内合成。
-
存在于突触前的末梢中,并且释放量足以引起突触后神经元或者目标器官上的激活。
-
当以合理的浓度外源性(exogenously)给药(作为药物)时,它会精确模拟内源性(endogenously)释放的递质的作用。
-
存在将其从作用部位移除的机制。
神经递质的清除一般存在以下形式:
-
被酶降解,比如乙酰胆碱(ACh)。
-
被转运蛋白主动回收(re-uptake),比如谷氨酸、GABA、甘氨酸等。
-
扩散或被细胞外肽酶降解(proteolysis),比如神经活性肽(neuroactive peptide)。这类递质作用距离较远并且分子较大,可能通过自然扩散稀释,或者被肽酶(peptidases)降解。
Lec 3. 初级视觉皮层 Primary Visual Cortex
视网膜
视觉的光感受包括视杆细胞和视锥细胞。视杆细胞负责暗视觉,无色觉,以多对一的方式对应到神经节上,敏感度低,与夜盲症相关;视锥细胞负责明视觉,有三种色觉,以一对一的方式对应到神经节上,敏感度高,与色盲相关。视锥细胞的三种色觉并不严格对应于三原色,而是具有短、中、长三种响应波长段。
视杆细胞中主要含有视紫红质,视紫红质中包含视蛋白和视黄醛。视黄醛在光的刺激下能够从顺式(11-顺视黄醛)转变为反式,从而引起神经过程。
感光的细胞以超极化为兴奋标志;它在暗处兴奋,在光亮处活动减弱。这与神经节细胞不同;在向神经节细胞传递的过程中,对于 ON/OFF 神经节的通路会分开。同时,视杆细胞之间存在横向的连接和侧向抑制。侧向抑制行为在视觉系统中普遍存在。
神经节/低级视觉处理
从感光到进入神经系统经过三层细胞,第一层为感光细胞(视锥细胞、视杆细胞),第二层为双极细胞,神经节位于最后一层。但是,在实际排布上,人类的视网膜的排布与光路是完全相反的;感光细胞位于三层细胞的最后面。光需要穿过神经节和双极细胞层,经历衰减之后才能够到达感光细胞层。除了在黄斑附近,神经节细胞向旁边倒开,光能够比较直接地传递到感光细胞层上。相对地,章鱼眼的感光结构排布与光路一致,因此不存在盲点。
神经节细胞具有 ON 和 OFF 两种类型,分别在更亮/更暗的时候产生更兴奋的反应。这种设计使得视觉系统在明处和暗处都能有较快的相应:相比使用神经元活动的减弱来表示光照变暗,使用 OFF 神经节的活动剧烈增强能够更快地向突触后的神经元传递这一变化。
神经节细胞具有感受野,大小定义为对应的光感受器的数量。对于一个神经节细胞而言,它的感受野可以被划分成中心区域和边缘区域。一个 ON 神经节细胞的响应如下:
-
只有中心受到强光照刺激时,产生兴奋的响应;
-
只有边缘受到强光照刺激时,产生兴奋的响应,但相比中心刺激响应更弱;
-
中心和边缘受到强光照刺激时,产生兴奋的响应,但相比中心刺激响应更弱;
-
中心受到光照刺激,边缘受到黑暗刺激时,产生兴奋的响应,并且比仅中心刺激更强。
OFF 神经节相似。这使得神经节细胞具有增强空间分辨度的能力,即在光照强度变化的区域产生更强的响应,而对均质的光照响应更弱。这事实上在对视觉信息进行基础的边缘检测操作,因此神经节具有低级视觉处理的功能。
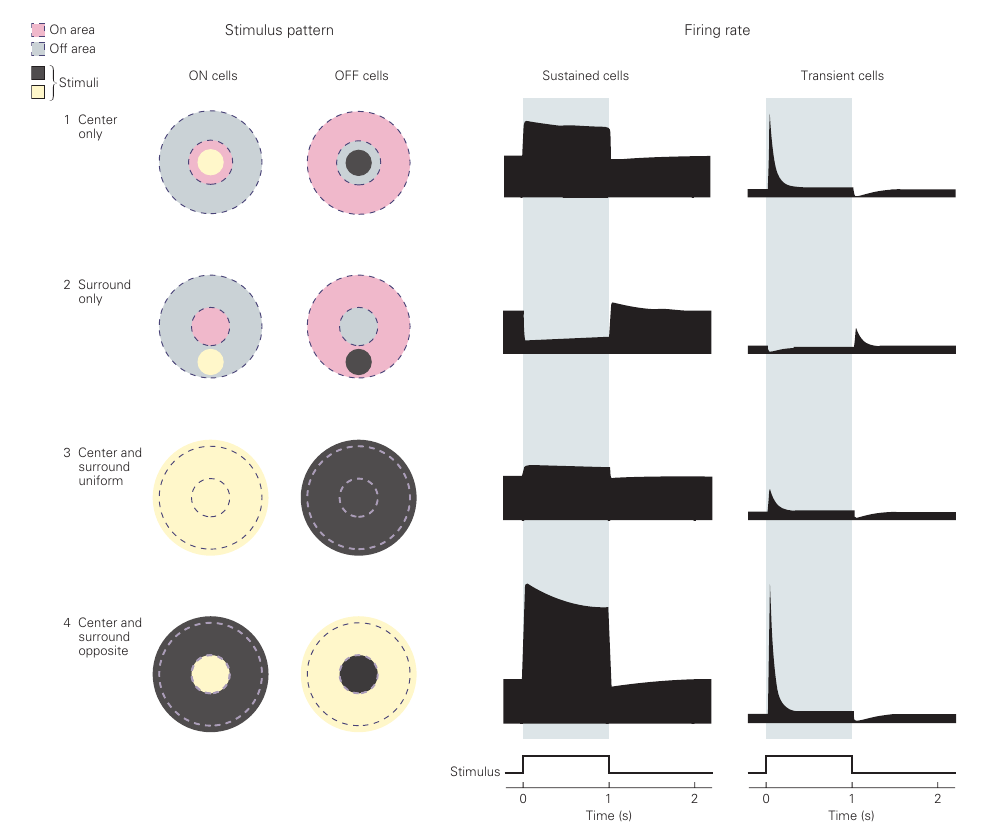
神经节细胞能够通过持续稳定发放动作电位,并通过发放的强度来表示响应的兴奋程度(sustained cells),也可以通过瞬时的响应增强来表示(transient cells)。总体来说,神经节细胞偏好对随时间变化的视觉信息进行响应。实验上,如果对固定的视网膜区域施加恒定的视觉刺激,视觉系统将在数秒内失去对它的响应。现实中并不会发生这样的事情,因为眼睛总是在进行扫视、漂移等操作,并不会真的盯着一个东西完全不动。
初级视觉皮层
初级视觉皮层具有方位选择性,是条状的感受野,即同心圆感受野的叠加。这里指的是初级视觉皮层的感受野,同心圆状的感受野来自外侧膝状体。原理如下。
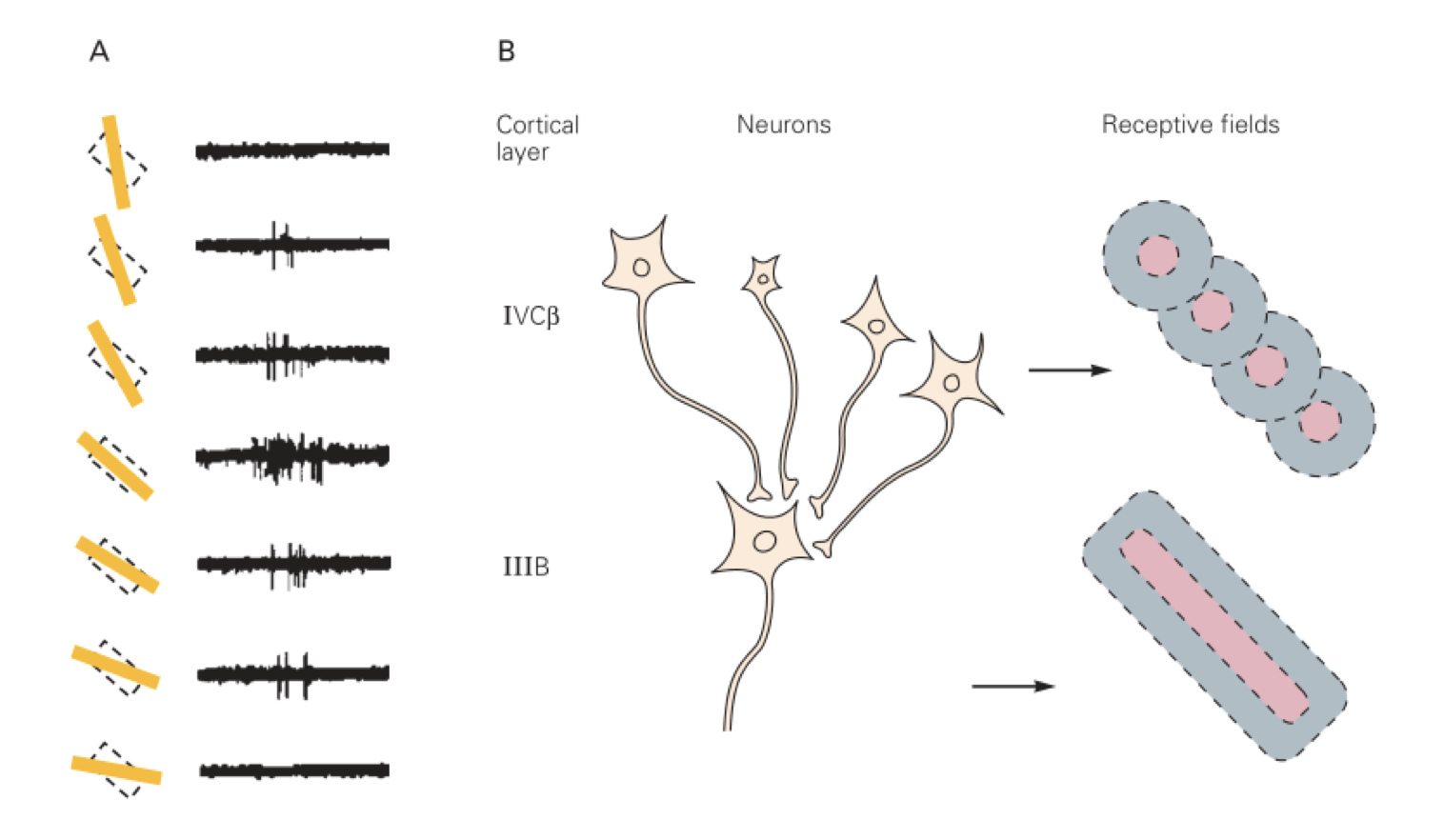
视觉系统的发育存在一个关键窗口期。在窗口期内,大脑的可塑性快速地发育成形。如果对幼年动物的视觉进行单眼剥夺,在发育成熟之后就会功能性失明。
Lec 4. 视觉物体感知 Visual Object Perception
视觉系统在经过视网膜之后,划分为背侧(dorsal)和腹侧(ventral)通路。背侧倾向于传递运动、方位、深度信息,腹侧通路倾向于处理颜色、形状等信息。
视觉信息通过神经节之后,进入外侧膝状体(LGN)。灵长类动物的外侧膝状体严格划分为六层,每个层对不同的信息敏感。第 1,4,6 层接收一侧眼的信息,第 2,3,5 层接收另一侧眼的信息。外侧膝状体后进入初级视皮层、次级视皮层等一系列视觉脑区,一部分最终达到前额叶,指导注意力,一部分最终到达海马体,形成视觉工作记忆。
初级视觉皮层中的简单神经元能够辨识特定位置的朝向信息,工作原理如上一节所示。简单神经元的感受野能够进一步叠加为复杂神经元,从而识别不同位置的方向信息。
初级视觉皮层形成柱状结构,同一个柱内的神经元具有相同的方向偏好性。灵长类动物的功能柱(方向柱)在不同方向区域的交界处形成风车状结构,风车状结构周围的方向偏好逐渐变化。一个完整的风车状结构称为一个超柱,超柱可以作为一个完整的功能单位,能够编码所有方向的信息。并非所有动物都能够形成柱状结构;小型啮齿类动物就没有观察到这种柱状结构。
方向柱不止存在于 V1,在其他皮层中也存在,且结构可能比风车状结构更复杂。不同朝向的功能柱不是均衡的,竖直和水平朝向对应的功能柱在 V4 中更多一些,因为人类接受的水平和竖直视觉刺激更多。
实验上的测量方法包括双光子成像、内源光信号成像(intrinsic optical imaging, ISOI)、功能性磁共振成像(fMRI)。从前至后,成像分辨率依次降低,成像范围依次扩大。
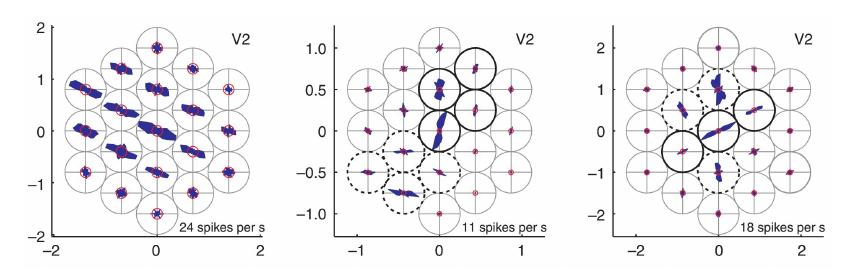
V1 的方向柱仅能编码简单的明暗条纹状朝向,对于更复杂的图案所编码的朝向信息(例如下图,右侧图片也可以编码水平/竖直朝向信息),V1 中没有相应的识别能力,但是在 V2 等区域中存在。

同时,在 V2 中,神经元的感受野也不再局限于同心圆状,而是通过抑制性的相互作用体现出对特定方向的选择性,形成更复杂的感受野。
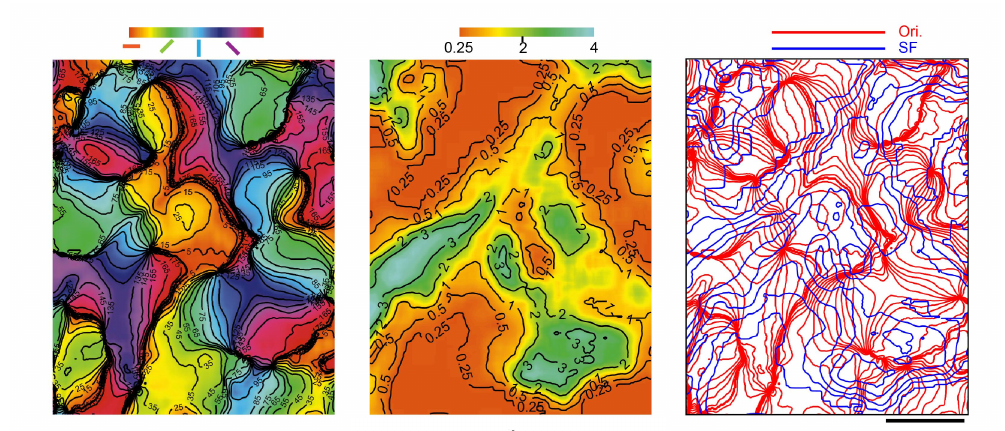
视觉皮层表现出高度的结构性。例如,连续的方向、位置、颜色、曲率刺激对应于连续的皮层区域。同时,对于不同的性质的信息,视觉皮层展现出的结构性倾向于互相正交,比如 V2 和 V4 中方向偏好的等势线和空间频率的等势线之间大多是正交的,即这些性质之间体现出一定的组合性。
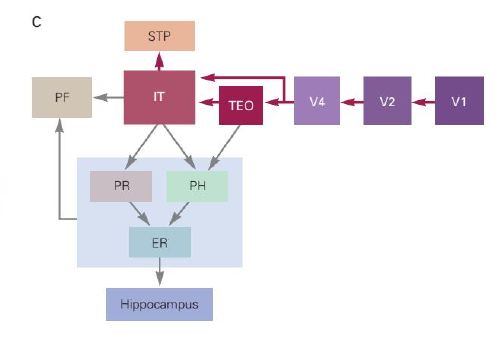
从简单的几何刺激到完整的物体识别(如面孔识别),其中的通路为何、如何进行信息的处理,其中的机制仍然是空白。两个与高级视觉处理相关的脑区位于 TEO(Temporal Occipital,枕叶与颞叶交界处)以及 IT(Inferotemporal,颞叶下缘)。TEO 受损将导致统觉性视觉失认(apperceptive agnosia),患者不能将物体复制地画下来,即虽然能够接收到视觉输入,发生了感知(perception)过程,但无法有效地将视觉输入的信息统合为一个整体,没有发生识别(recognition)过程。IT 受损将导致联想性视觉失认(associative visual agnosia),患者虽然能够将物体画出来,却不能够说出这个物体究竟是什么,及无法将视觉的认知与语义联系起来。
Associative visual agnosia refers to a subtype of visual agnosia, which was labeled by Lissauer (1890), as an inability to connect the visual percept (mental representation of something being perceived through the senses) with its related semantic information stored in memory, such as, its name, use, and description.
从视网膜到脑区的映射是完全倒置的。左侧视野映射到右脑,右侧视野映射到左脑;上半视野映射到下半部分,下半视野映射到上半部分。
颜色感知
视锥细胞仅在黄斑区域集中分布,其他区域分布得较少。因此,人类只能在中心视野中比较敏感地分辨颜色。
人类对颜色的感知严重依赖于相对外周的环境,而不是绝对的波长。视觉系统的感受野决定了这种感知:感受野边缘的响应会影响到位于感受野中心的响应。
Lec 5. 运动和空间感知 Motion and Spatial Perception
运动处理
运动和空间感觉由背侧通路完成,经过 V1 和 V2 之后进入 V3,之后进入 MT(V5),进入顶叶。顶叶中包含本体感觉的形成,视觉信息能够告诉顶叶当前个体在空间中的什么位置,以及其他物体相对它的运动状态。实际上很多信息是直接从 V1 直接传入 MT 的,因为对运动信息的处理需要很快。
虽然背侧和腹侧通路都经过 LGN、V1 和 V2,但是在细分的经过区域上不尽相同。在 LGN 的六层结构中,含 M 细胞,与深度和运动视觉相关;P 细胞,与颜色和边界有关;K 细胞,与颜色有关。
实验上,神经活动的物理连接有两种类型的示踪剂:顺行(anterograde)示踪剂,从胞体向轴突传播,图像呈网状,能够判断出信息传递的下一级位置;逆行(retrograde)示踪剂,从轴突末端向胞体传播,图像呈点状,能够判断出信息的来源位置。
MT 是运动处理中的高级脑区,在进化中相对保守,很多生物中都存在对应区域。与物体感知相比,运动处理相对应的脑区相对都比较小,同样是由于需要较快的处理。在实验上,MT 是通过髓鞘染色界定的,因为 MT 中髓鞘含量很高,能够被深染色。髓鞘含量高的原因同样是需要快速的传导。
各个视觉脑区中都有一定的神经元具有运动方向选择性,尤其是 MT。
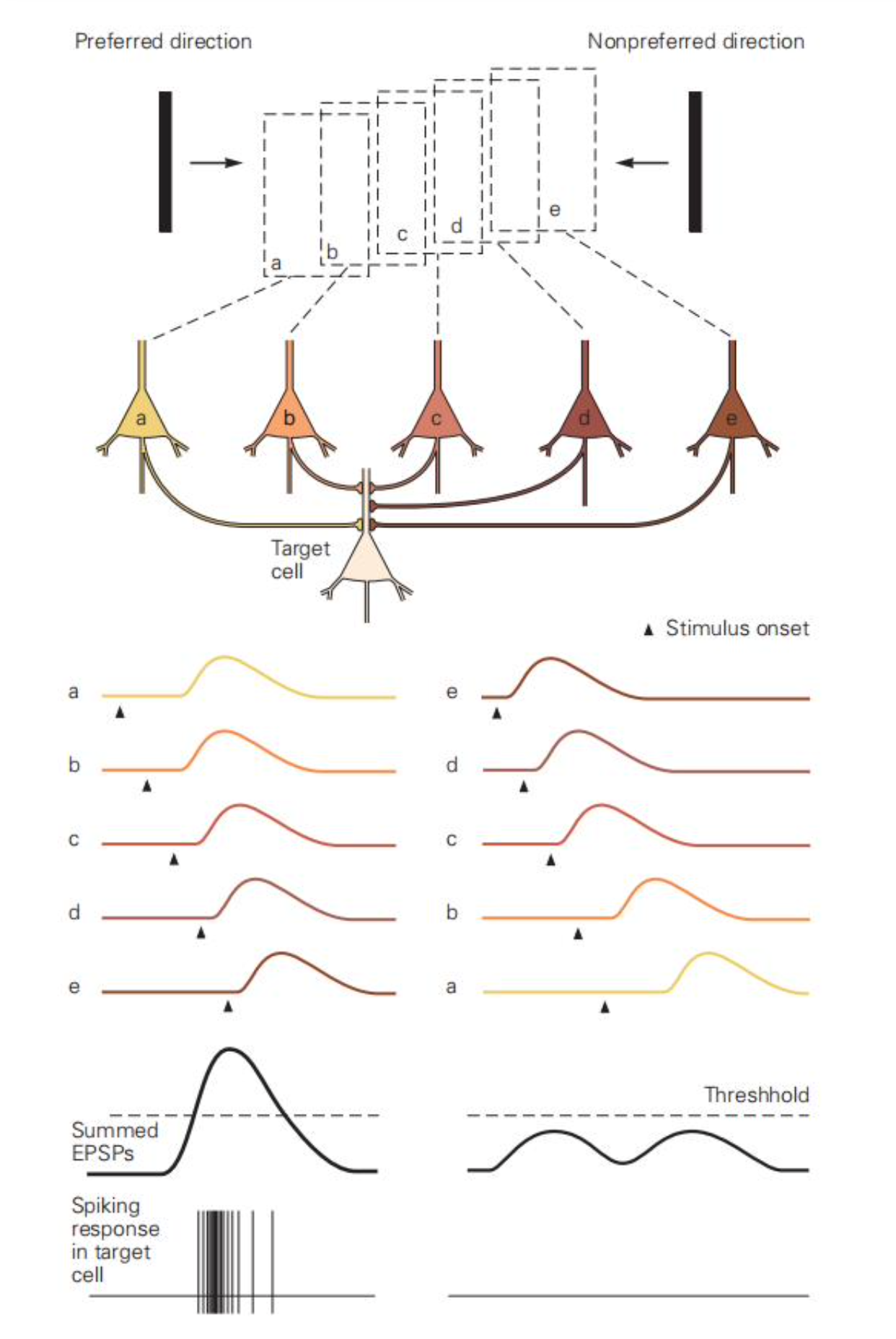
对于方向选择性的形成,存在一种假说:一系列感受野连续的神经元向同一个神经元输出(类似朝向选择性形成),且他们对刺激的响应有一定的延迟,且延迟依次变短。如果刺激先进入延迟长的神经元的感受野,后进入延迟短的神经元的感受野,它们的响应会由于延迟叠加在一起并超过阈值,从而引发对特定方向的响应。其他运动方向的刺激由于不能在延迟上恰好符合,所以只能引发阈值下的行为。实验仅验证确实存在感受野连续的神经元,但尚未验证存在同时满足感受野和响应延迟连续的神经元。
在对运动方向具有选择性的神经元中,一部分神经元进一步地对速度具有选择性。
MT 中的方向选择性确实影响到人对运动的感知。对 MT 中具有运动方向选择性的神经元进行刺激,将会影响到被试对运动方向的判断。
在更加复杂的运动方向模式中,也存在选择性的响应。比如,对于两个向不同方向运动的光栅,V1 中存在神经元响应于合成运动的运动分量,MT 中存在神经元响应于整体的合成运动方向。对于合成运动方向的感知是需要一段时间内逐渐感知形成的。
除了简单的单方向运动,还存在特定的神经元能够响应顺时针、逆时针、轴向放大、轴向缩小等等复杂运动模式。这些响应模式的形成过程尚不清楚。
运动信息会进一步传递到前额叶,用于对眼球的运动进行反馈控制。
朝向选择性是天生的;运动方向选择性是后天习得的。
空间定位
灵长类动物的高级空间感知能力是由双眼视觉带来的。灵长类动物的双眼均长在面部前方,而非两侧,这虽然使得视场较小,但双眼的视觉范围有较大的重叠。人的双眼有相同的注视点,但左右眼看到的图像并不相同,存在一定的左右偏移。基于这样的微小偏移,视觉系统就能够建立立体视觉。这实际上也是立体镜以及立体电影的原理,即让双眼看到不同的图像。
LGN 和 V1 IV 层均不存在双眼视觉相关的神经元,因为它们只接受单眼的视觉输入。在 V1 IV 层向 II、III 层传递的过程中,才会出现同时接受双眼视觉的神经元。V1 中没有形成双眼视觉相关的功能柱,在 V2 中才形成。
除了借由双眼形成的立体视觉,借助物体本身的运动也可以形成单眼的立体视觉,即不同视觉深度的物体运动的速度不同,即视场中一定的速度梯度。一种可能的机制是一些神经元通过非经典感受野对特定方向的速度梯度响应。
Lec 6. 注意和工作记忆 Attention and Working Memory
注意是选择性地集中于信息的离散部分的认知过程。
注意力分为自下而上的和自上而下的。自下而上的注意力中,在场景的刺激中,大部分的刺激是同质的,只有少部分的刺激非常特殊,即使我们没有去特别关注,注意力也会被自然地引导到异常值(salient)所在的位置。自下而上的注意力是由刺激引导的、并行的、快速的,处理的速度与刺激的数量无关。自上而下的注意力中,场景中的刺激都是异质的,我们可能被要求去注意其中符合特征的某一个。我们必须逐一地检查场景中的刺激,因此自上而下的注意力是主观引导的、串行的、缓慢的,处理的速度与刺激的数量有关。
注意的对象可以是比较广泛的。比如,注意某一个特定的空间位置,注意具有某种特征(颜色、质地)的对象,或者注意某一个特殊的物体(比如脸)。
自底向上的注意力的神经基础被认为是周围抑制(surround suppression/contextual modulation),即感受野内的响应受到同质刺激的抑制和异质刺激的增强。自底向上的行为即使在麻醉动物中也能观察到。自上向下的注意力由更高级的脑区控制。
空间注意的经典实验任务是 Posner Cueing Task。任务中,会给出一个 cue,比如箭头,暗示接下来的刺激的位置,之后刺激会出现在指示的位置,或者出现在与指示不符的位置。被试的反应速度会发生响应的变化。注意会增强神经响应的敏感性;比如,在有空间注意的情况下,神经元的朝向选择性会进一步增强,体现为 tuning curve 上的乘性缩放。
注意不仅影响注意的目标,也会影响周围区域对应的神经响应。
意识 consciousness
意识的一类研究方法是通过双义(bistable)的图像,或者双眼竞争视觉(Binocular Rivalry)。在双眼输入不同的视觉输入的情况下,被试会看到不稳定的图像,尽管刺激是恒定的。
工作记忆 Working Memory
一个早期的工作记忆是 Baddeley 工作记忆模型。他假定工作记忆中存在
-
Phonological Loop,以类似声音的形式记忆。比如,当一个人记忆电话号码的时候。
-
Visualspatial Sketch Pad,视觉工作记忆。
-
Episodic Buffer,事件记忆。
工作记忆的经典任务是延迟响应,即被试先被展示一些信息,然后经过一段延迟的掩盖,最后根据记忆的信息完成一些任务,其中可能包含一些额外的推理。
空间工作记忆位于背外侧前额叶皮层(dorsolateral prefrontal cortex),即前额叶的上半部分。相对地,腹外侧前额叶皮层(ventrolateral prefrontal cortex)的活动反应了对象(object)工作记忆的保持。对象工作记忆指的是针对特定物件的记忆;比如,先向被试展示一个物件,然后在遮蔽一段时间之后让被试从一堆物件里面选出先前见到的那个。相对地,空间工作记忆的例子是在两个地方分别放置一个物体,并在遮蔽之后让被试指出指定物件的位置。一种假说认为视觉信息通过背侧通路进入 DLPFC,通过腹侧通路进入 VLPFC。但这种观点仍存在质疑;实验上,工作记忆可能表现出分布式的性质,在感知、顶叶、颞叶、前额叶等区域都能够观察到类似工作记忆的活动。
Lec 7. 听觉系统 Auditory System
听觉系统相关疾病:耳聋(先天性/随年龄衰老)、耳鸣(长期暴露在高噪声环境中,持续听到实际上并没有的声音)、孤独症、精神分裂
听觉系统与视觉不同,是一个天然含时间的过程,并且可以接受全方位的输入。另外,听觉系统能够接收来自自己的声音,与自己的发声形成强烈的反馈;比如,当自己在戴着耳机时,说话会特别大声。此外,听觉系统的进入皮层之前有更长的通路。
人对声音频率和声压(sound press level)的感知是对数尺度的。
听觉外周 Auditory Periphery
外耳具有声音聚拢、选择性放大和定位的功能。外耳道具有一定的共振性质,对于特定频段的声音具有放大作用。此外,对于不同位置的声音来源,外耳道产生的频段增益会有所不同,因此能够产生一定的声源方位信息。中耳通过听小骨将声音的衰减进行放大弥补,内耳将会把机械振动转换为电信号。信号的转换由毛细胞完成。毛细胞的上端有纤毛,受到机械振动时,纤毛会发生摆动,同时上面的离子通道打开,引起 Ca2+ 内流。
在经过毛细胞转换为电信号之后,听觉信号进入听神经传播。听神经表现出群体编码的性质;不同的听神经纤维有不同的偏好频率(Best Frequency,BF),或者称为特征频率,从而携带不同频段的信息。
听觉中枢
听觉通路:毛细胞(hair cell)-听神经(auditory nerve)-耳蜗核(cochlear nuclei)-上橄榄核(superior olive)-下丘(inferior colliculus)-内侧膝状体(medial geniculate body)-听觉中枢(auditory cortex)。整个听觉通路从脑干进入中脑,然后是丘脑,最后进入到听觉皮层中。听觉系统既有上行通路,也有向下反馈的下行通路。
声源定位也是通过双耳听觉实现的,即不同的声源进入左右耳的时间和强度会有所不同,这个过程发生于上橄榄核,其中中央核(medial superior olive)编码了时间差,不同时刻到达中央核的刺激时间差会被清晰地记忆;外侧核(lateral superior olive)编码了强度差,从同侧进入的刺激是兴奋性的,但对侧进入的刺激会被转换为一致性的,从而进一步放大强度差距。
听觉皮层 Auditory Cortex
听觉相关的皮层位于颞叶。对于人类来说,人类的听觉皮层在颞叶的沟中,基本不会外露。刺激耳蜗的不同位置,将会得到初级听觉皮层不同区域的响应,称为 tonotopic map(频率拓扑)。初级听觉皮层(A1)的 tonotopic map 是条带状的,同一个条带中的神经元具有相似的 BF。听觉中枢的核心区域根据 tonotopic map 的变化趋势,又划分为 A1、R 和 RT。
在核心区域外侧,有 belt 区域环绕核心区域。belt 外侧还有 parabelt。parabelt 一般不激活,只有在完成具有奖励的特定听觉任务时会被激活,并向前额叶传递信号。
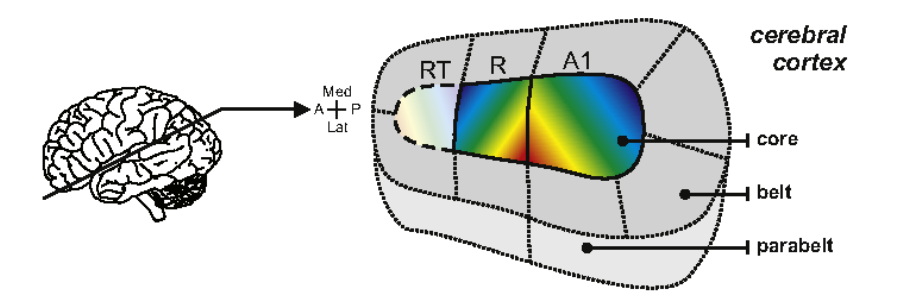
根据功能的不同,听觉信号向听觉皮层传导的通路由被划分为丘系通路(lemniscal pathway)和非丘系通路(non-lemniscal pathway)。丘系通路有明显的频率拓扑,且忠实地传递听觉信号;非丘系通路没有明显的频率拓扑,并且涉及到多感觉的整合。
在经过听觉皮层之后,与视觉系统类似,分为 what 和 where 通路,分别通过腹侧和背侧通路最终到达前额叶。
在实验上,对听觉中枢的记录通常要求在清醒的动物中记录。在麻醉的动物中,听觉中枢的神经元会非常沉默。
中枢系统中的神经元对声音刺激有强烈的偏好性。越靠近中枢系统,偏好性越高。
Lec 8-9. 杂谈(?)
电生理记录
电生理记录的基本单位是动作电位。
局部场电位(Local Field Potential, LFP)是胞外记录,反应的是一群神经元同步活动的信息。如果有活跃的发放或者超极化的活动,LFP也会发生相应的变化。EEG则范围更大,同理。
在电极插入时,可能会同时记录到周边的多个神经元的发放信号。为了从记录中区分不同的神经元来源,需要同时插入多个电极,利用位置差进行区分。
对于细胞外记录到的原始数据进行滤波,高频段记录的是脉冲的发放,低频段记录的是平均场电位、alpha 波和 gamma 波的信息。对于脉冲的数据,通常会观察发放频率或者脉冲间的间隔时间分布等等。
在没有其他植入的情况下,光成像和超声成像的主要方法还是依靠血红蛋白的氧合和脱氧特性。除了血红蛋白之外,最常用的就是荧光蛋白(GFP)和电镜技术。
睡眠
在最极端的情况下,一直没有睡眠会导致身体各项机能的崩溃,长期的睡眠剥夺最终会导致死亡。自然界睡眠最多的是蝙蝠类,最少的是牛马一类容易被捕食的生物。
在定义上,睡眠是自然的、容易逆转的、失去自主意识的状态,伴随梦和大脑活动的变化。人类的睡眠包括快速眼动期和深度睡眠。
睡眠最主要地受到光照的节律影响。次要地,也受到内源性的调控,比如午睡。
睡眠受到节律影响可以在基因表达的层面上找到证据。
关于睡眠的假说:
-
睡眠能够积蓄能量(冬眠);
-
能够促进学习和可塑性的发生;
-
能够清除代谢产物,恢复代谢的状态。
光遗传学 Optogenetic
光遗传学起源于对感光细胞的研究。使用光调控神经元的活动在上个世纪就已经出现,因为光具有足够的空间分辨率和时间分辨率,能够做到调控单个神经元在神经活动尺度上的行为。基本原理是将光敏感的离子通道蛋白转移到神经元中表达。
memory
记忆分为外显的(explicit)和内隐的(implicit)。外显的记忆能够用显式地表达出来,比如记住一串数字,然后复述;内隐的记忆包括习惯、肌肉记忆等等。对海马区域的切除不会影响隐式记忆的形成。
对于记忆的形成,一种假说是记忆存储在突触连接的模式中,即由神经元之间连接与否的模式决定。另一种假说是记忆通过突触连接的强度存储。神经科学上,连接的强度体现为给予刺激时能够引起更强的 EPSP,或称为长时程增强(Long Term Potentiation,LTP)。LTP 和 LTD 构成 STDP,即取决于突触前后刺激发生的时间,改变 EPSP 的响应。除此之外,在海马区域,取决于两个突触前输入到达之间的时刻匹配程度,也会产生 EPSP 的变化,称为 input-timing-dependent plasticity(ITDP)。
突触并不是单向的连接。突触后接受受体之后,会向突触前释放递质,从而反过来调控突触前的行为。
现在对于海马体的认知是:海马体是短期记忆的存储地点,皮层到海马之间存在显著的连接。在执行任务的过程中,需要长期储存的记忆逐渐固化到皮层区域的连接中,海马体继续存储后续的短期记忆。
体感皮层和运动控制
身体感觉的通路起始于连接身体表面的感觉神经元,随后传入脊髓、脑干,然后进入丘脑,最后到达中央后回的体感皮层。脊髓的前角包含感觉神经元,后角包含运动神经元。在进入丘脑之后,处理区域都呈现出与身体部位对应的关系,不同部位的信号平行传输,但互相之间有交互和多感觉的整合。不同的感觉(温度、痛觉等)是分开传输的,受到刺激时发放的模式也不同,即编码方式不同。
Lec 10. 核磁共振原理
核磁共振的主要结构包括主磁体、梯度系统和射频系统。主磁体在MRI机器中产生高强度的均匀磁场,梯度系统由于进行空间定位,射频系统包括发射和接收装置,提供用于共振的波,接收装置用于接收共振导致的信号。
核磁共振在医学上可以用于结构成像和功能成像,结构成像包括血管成像等等。
成像原理
核磁共振主要依赖于原子的自旋性质。当原子核中的质子数量和中子数量不同时为偶数时,原子表现出一定的自旋方向。MRI成像依靠的主要是氢原子,因为人体内含有大量的水,有大量的氢原子可以用于MRI成像。另外,C-13和磷原子等也可以用于MRI成像。
在施加一定的外部磁场之后,氢原子的自旋方向将会由杂乱的排列回到统一的状态。其中,低能级的氢原子指向一个自旋方向,高能级的氢原子指向另一个自旋方向。氢原子的自旋方向并非处于静止状态,而是根据一定的频率产生进动(参考天体力学/陀螺仪原理)。初始状态下,氢原子之间的进动相位不同步,宏观的总磁通量方向仍然指向低能级的自旋方向,称为纵向磁通量。
在发生共振时,射频系统发出与氢原子进动频率相同的信号。这个信号一方面使得氢原子从低能级跃迁到高能级,并控制处于低能级和高能级的氢原子数量大致相同,从而在纵向上发生抵消。另一方面,这个信号使得氢原子的进动相位之间发生同步,即共振,从而在横向上形成一个按照进动频率进行旋转的横向磁通量。射频系统的接收装置是一个线圈,在横向磁通量不断穿过这个线圈的过程中,线圈中将产生感应电流(楞次定律),从而捕捉到共振信号。这个感应电流将会呈现三角函数震荡的形式。
在撤去射频信号之后,氢原子的状态将会逐渐回到原始的状态。一方面,高能级的氢原子逐渐回到低能级,使得纵向磁通量逐渐恢复。纵向磁通量的恢复遵循负指数规律,特征时间(即恢复到原始纵向磁通量的63%)的时间称为T1。纵向磁通量的恢复速度取决于周围组织的热性质,因此能够用于区分不同的部位。另一方面,氢原子之间的进动将会逐渐失去相位同步,从而使得横向磁通量衰减。这个衰减将会引起感应电流在震动幅值上的衰减,这个衰减也符合负指数规律(与上面不同,指的是和的区别),它的特征时间(即衰减到原始值的37%)称为T2。这个过程中产生的信号被称为自由感应衰减信号(free induction decay, FID),是MRI的一个主要信号依据。这个过程被称为横向弛豫/纵向弛豫。
自由感应衰减信号可以直接用于成像。不同组织的T2不同,在给予射频信号之后的一段时间中,一些组织的信号衰减多,一些组织的信号衰减少,从而形成不同的成像亮度。这种方式称为T2加权成像。
T1加权成像指的是在纵向磁通量恢复的不同阶段再度给予射频信号。一些组织纵向磁通量恢复得快,此时给予射频信号将会导致再度引发一个较强的横向磁通量,反之则引发得比较小(不太理解,但反正是这么回事x)。成像结果与T2加权成像相反。结构相成像主要是T1加权成像。
质子密度加权成像指的是在给予刺激之后,立刻进行记录(TE=0)。此时得到的成像表示了不同组织中的氢原子密度。
TR指的是两次给予射频信号之间的时间间隔(Relaxation),TE指的是在射频信号发出之后到进行记录之间的时间间隔(Echo)。对于T2成像,TR长,TE也长。对于T1成像,TR短,TE也短。
定位原理
在MRI系统中,梯度系统用于空间定位。平行于主磁场的方向标记为z轴;梯度系统在z方向上施加一个额外的磁场,从而使得磁场强度沿着z轴发生线性的变化。当给予射频信号时,只有磁场强度满足特定值的z位置才会发生共振现象,从而选定指定z位置处的xy平面,这一过程被称为选层。为了进一步确定xy平面上的位置,在信号采集前和信号采集时分别会在xy方向上施加磁场强度梯度,从而改变共振的相位(采集前)和频率(采集时),最终利用傅立叶方法进行空间定位。细节略,看不懂。
一个经典的MRI SE信号序列如下所示。虚线之间的时间为0.5TE。90度射频即上述提到的共振射频信号,180度脉冲是一个额外的采集手段。实验中,由于人体等其他物体对磁场强度的影响,横向磁场的衰减速度往往远快于T2规定的速度。180度脉冲用于引起一个自旋回波,便于信号采集。
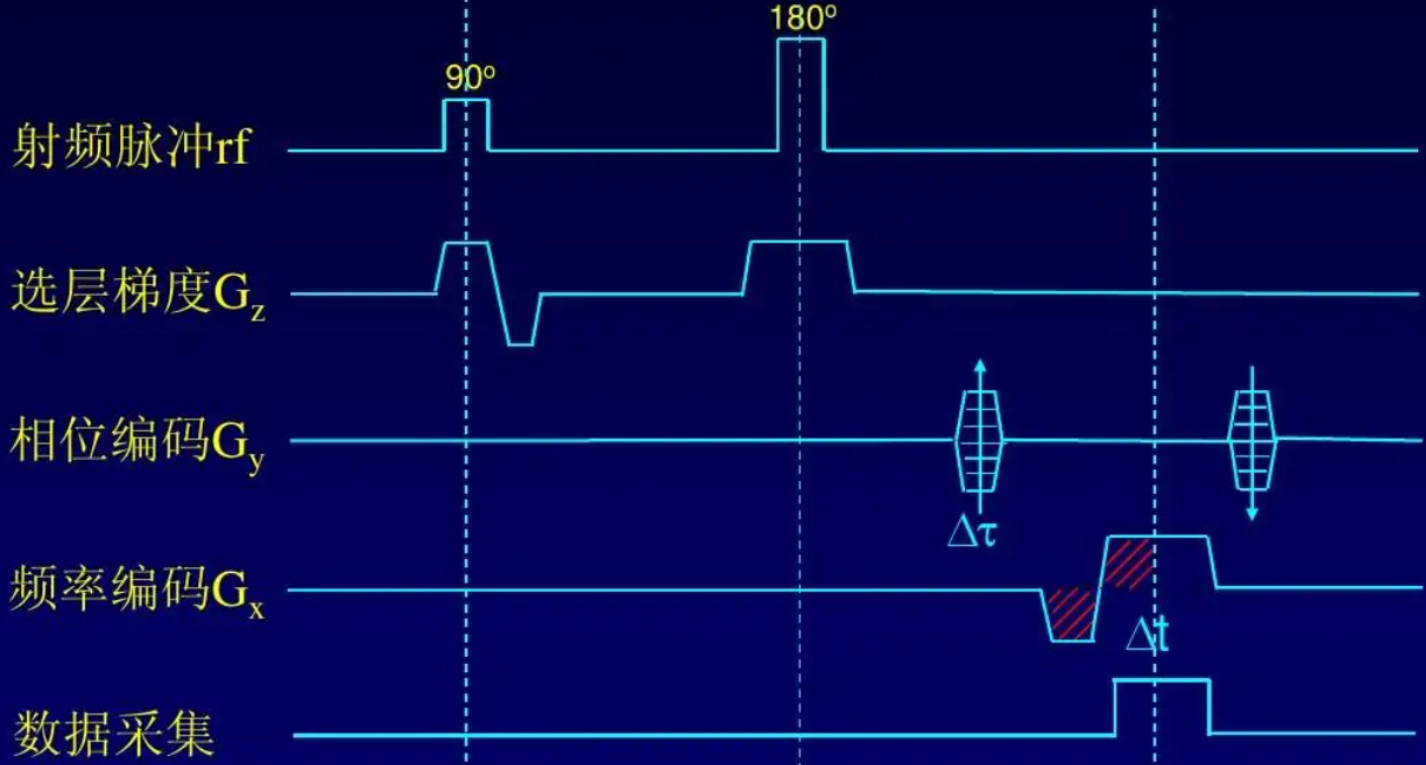
fMRI
BOLD(Blood Oxygen Level Dependent signal)是功能性核磁共振成像的主要手段。在神经活动活跃的区域,氧合血红蛋白的成分会增加。血红蛋白在不同的状态下会改变顺磁性和抗磁性,从而产生不同的磁环境,使得氢原子的T2延长,最终影响到成像的结果。
BOLD的原理依然存在一定的争议,因为血氧的变化并不一定意味着神经活动的变化。
fMRI实验设计
fMRI就是现代颅相学,即试图将某种功能对应到某个脑区域。
fMRI实验的主要逻辑是对比试验/减法逻辑,即对比两个刺激所激活的区域,有差异的区域代表了两个刺激之间的差异所激活的区域。
-
Block Design。指的是给予一个刺激,持续一段时间,然后切换成另外的刺激。block design的主要设计因素是单个刺激的持续时间,时间太短不能观察到显著的活动变化。
-
ER Design。指的是给予一个短刺激,然后静默一段长时间或者短时间,然后再进行短刺激。可以用于研究意外事件。
fMRI adaptation指的是在给予重复的刺激时,fMRI响应可能会减弱。如果在一个体素中既含有对A敏感的神经元又含有对B敏感的神经元,整个体素的平均响应可能不会体现出对A或者B的敏感性。此时,按序列呈现AA刺激或者AB刺激,前者应该呈现出更弱的反应,从而确定体素中可能含有对A敏感的神经元。fMRI adaptation的底层原理尚不清楚。
Lec 11. 情绪/边缘系统 Emotion & Limbic System
情绪与情感密切相关。前者表现为短期的反应,后者表现为长期的效应。情绪同时与其他高级功能相关;比如,当情绪处于亢奋状态时,对应产生的记忆也会更加深刻。
人类情绪最集中的部位是眼睛和眉毛。单纯通过观察眼睛和眉毛的状态,就能够编码非常多的情绪状态。
关于情绪的一个早期理论(James-Lange)是,生理变化是情绪的诱因。外界刺激导致生理状态的改变,这种改变影响到大脑,大脑从而产生对应的情绪反应。比如,饮酒导致心跳放缓,进而导致放松的情绪状态。但这种理论存在一些缺陷;有时候不需要明显的外界刺激,也会产生情绪体验,并且某些生理变化也不直接与情绪相关,比如瞳孔收缩。另一种理论(Cannon-Bard)认为,外界刺激传入到大脑皮层之后,产生了某些特定的情绪响应,这些响应进一步改变了生理状态。此外,情绪与过往的经历有关,比如根据人的过往经历,可能会对某些场景产生特定的情绪或者放大情绪(杯弓蛇影)。因此,也有理论认为情绪是皮层对过往经历的投射。比如,在 The Little Albert 实验中,幼儿会联系锤子的敲击和小鼠,从而在看到小鼠时产生恐惧情绪。
现代的情绪理论认为,外界刺激同时进入皮层和下丘脑,下丘脑能够产生较快的响应,皮层能够产生更高级的活动(比如根据过往经历进行响应),并进一步调控下丘脑的行为。下丘脑的活动调控脑干等区域,调节激素活动,从而形成生理的响应。
边缘系统位于皮层以下,扣带回和下丘脑以上。广义上,下丘脑等与情绪相关的核团都包括在边缘系统中,包括杏仁核(amygdala),主管恐惧;扣带回,含有镜像神经元,主管共情能力等。
愤怒行为产生于中缝背核(Dorsal Raphe)。中缝背核分泌5-羟色氨酸,调控下丘脑的行为,从而产生愤怒和攻击性的响应。
快乐与隔核、伏隔核和腹侧背盖核相关,且与特定的神经递质相关,即多巴胺。
前额叶与更加复杂的社会行为有关。例如,前额叶受损的患者在面临电车难题时,会毫不犹豫地选择拉下拉杆,表现出一定社会道德的缺失。
Lec 12. 小脑 Cerebellum
小脑占据整个脑体积的10%,但是神经元数量非常之多,甚至比大脑要多。小脑具有很规则的结构。传统的观点认为小脑主要负责运动控制,但实际上小脑负责了广泛的调节功能,如果小脑出现一定的障碍,情绪等功能也会出现障碍。
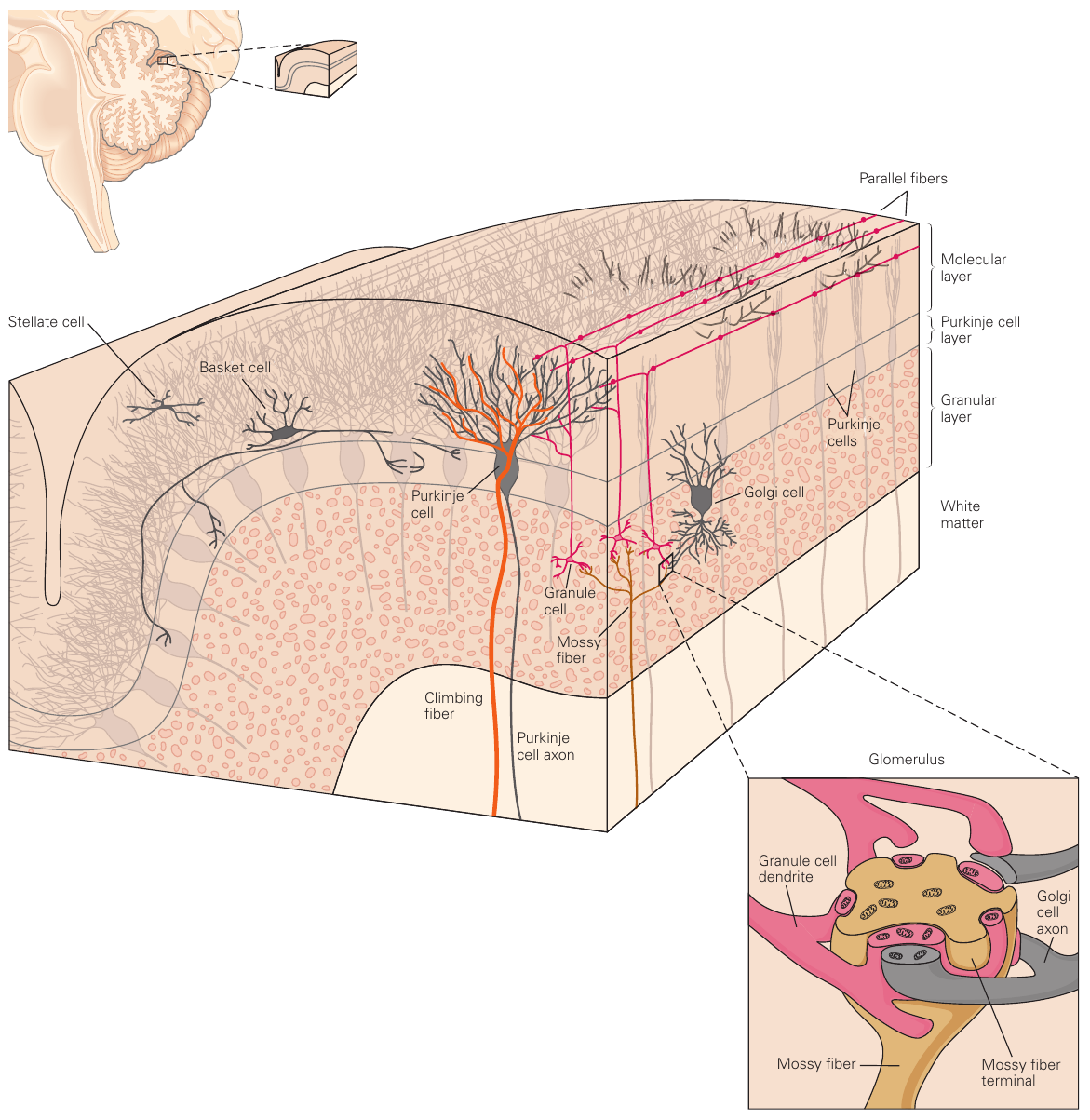
小脑上有大量的沟和回,从后至前划分为绒球小结叶(flocculonodular lobe),小脑后叶(posterior lobe)和小脑前叶(anterior lobe)。从左右两侧向中间划分为hemisphere,intermediate part以及vermis。小脑内有四对核团,包括顶核、球状核、拴状核和齿状核。小脑通过三对小脑脚与大脑连接,连接非常松散,解剖的时候会掉下来。
绒球小结叶,或称前庭小脑(vestibulocerebellum),是小脑中最古老的部分,最早出现在鱼身上。绒球小结叶与前庭相互连接,接受来自前庭和视觉系统的输入,参与平衡控制、前庭眼球反射和眼动等过程。
脊髓小脑(spinocerebellum)包括中央的vermis和intermediate part。它接受来自脊髓的躯体感觉和本体感觉(即个体目前的位置等信息)输入。
皮层小脑(cerebrocerebellum)是指hemisphere部分,主要与大脑皮层连接。与运动规划相关。该部分如果受到损伤,会影响到大脑的认知功能。
小脑的输入输出基本遵循从哪里来到哪里去的原则。前庭小脑的通路独立于小脑之外,不经过小脑内部核团,来自前庭并最终返回前庭;vermis的输入先进入顶核,intermediate part的输入先进入球状核和拴状核,然后传输到小脑皮层,最终回到脊髓;hemisphere的输入先进入齿状核,然后经过皮层回到大脑。
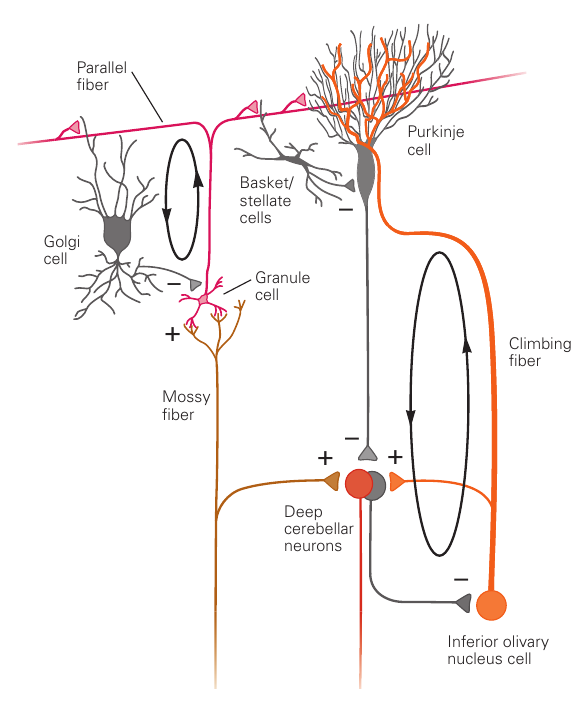
小脑自外向内分为molecular layer,Purkinje cell layer和granule layer。颗粒细胞层细胞数量非常多,是兴奋性的;其他层的细胞大多是抑制性的。小脑的输出仅来自于浦肯野细胞,且是抑制性的。小脑的输入主要来自于两种神经纤维。mossy fiber连接来自脑桥的输入,climbing fiber连接来自下橄榄核的输入。mossy fiber从颗粒细胞的树突进入小脑皮层,并传导到molecular layer。颗粒细胞的末端在molecular layer形成平行的神经纤维束,称为parallel fiber。单个parallel fiber连接大量的浦肯野细胞末梢,从而产生广范围的影响(类似矩阵作用)。climbing fiber沿着浦肯野细胞的方向进行爬升,并包裹住浦肯野细胞的末梢,直接进行接触。一个浦肯野细胞的末梢仅接受一条climbing fiber的输入,但是一条climbing fiber可以影响多个浦肯野细胞。浦肯野细胞的输出频率非常高,在动物处于静止状态时就会以100Hz左右的频率稳定发放;在发生活跃的运动时,发放频率则会达到数百Hz。
小脑皮层完整的环路如下。其中包括一些循环的控制单元。
当小脑出现功能障碍后,最主要地会出现运动相关的问题,也可能出现其他功能的障碍。包括肌张力降低,体现为膝跳反应时会跳很多下;立行不能、共济失调,即不能站立或者步幅步态不协调,很难开始运动。基本上所有无意识的运动都会受到影响,尤其是多关节协调的运动。
小脑和大脑的绝大部分区域都不一样,大脑是对侧控制的,但小脑是同侧控制的。如果左侧小脑受损,影响的就是左侧的运动。小脑的浦肯野细胞和内部核团的放电率一般非常高,用于控制主动运动。小脑对运动的控制是预测性的/前馈性的,区别于反馈控制。
Lec 13. 核磁共振数据分析
预处理
对于结构相数据,原始数据需要映射到标准空间。目前两种标准空间(Talairach和MNI)都是西方人的脑模型,事实上东方人的脑和西方人的脑模型还是有一定的区别。
对于功能相数据,需要进行:
-
slice time correction。这指的是一次全脑信号扫描的不同切片实际上是在不同的时间进行的,反应的不是同一个时间点的状态。需要根据同一个切片不同时间点的状态进行插值。
-
3D motion correction。指的是被试的头部运动导致的矫正。被试的头部运动会导致记录位置不准确,同时也会对磁场产生干扰。
-
distortion correction。主要是由磁场不均匀造成的图像变形。
-
temporal filtering。主要用于消除基线的信号漂移、心跳和呼吸带来的干扰等。
-
spatial smoothing。降低一定的空间噪声,但是同时也会降低有效的空间分辨率。通过卷积等方式实现。
-
spatial transformation。指的是将功能像与结构像进行对齐。
数据分析
对于单个体素而言,fMRI最终得到的结果是一个时间序列。由于我们知道这个时间序列的每个时刻实际上给予了什么刺激,基础的数据分析的过程就是将这个时间序列上的相应与每个时刻对应的刺激标签进行线性回归,然后评估相关性。这种方法被称为广义的线性方法(Generalized Linear Model, GLM)。
GLM是单体素分析的方法,MVPA(Multi Voxel Pattern Analysis)指的是多体素的分析方法。多体素是指将不同的体素的激活情况拼接成一个特征向量,然后使用ML的方法进行处理。常见的方法是经典SVM;比如,要评估一个区域是否对AB两种视觉模式可分,每个试次中会获得这个区域的一个激活特征模式,然后利用这个模式训练SVM,评估SVM的可分性。
更高级一些的ML方法假设每个体素对应于一些特征,比如在训练阶段给被试看一定的图像,推理阶段通过神经活动重建图像的某些特征。
另一类方法是基于相似度矩阵的方法。比如,在给予被试不同刺激时,被试的脑区会给予相同或不同的相应模式,这些模式之间可以形成相似度矩阵。这种方法也可以判断一个脑区是否对某种输入特征敏感。相似度矩阵可以处理高度灵活的两种输入;比如,可以比较两个脑区的活动,从而判断两个脑区之间的连接强度;可以比较跨物种的脑区;可以比较生物的脑区与人工神经网络之间的活动。相似度矩阵可以视为距离矩阵,从而进一步可视化为地图或者层次聚类的形式。
Comments NOTHING